這是一個很好的問題,而且是不能很簡單的解釋的問題。我嘗試提供幾個方面的視角,並歡迎大家提出意見以供討論。
1. 視覺感知原理:視覺編碼
要回答題主的問題,我們可以先考慮跟高層面的 視覺的感知 ,從人腦視覺編碼方式說起。
1.1 視覺通路 (Visual Pathway)
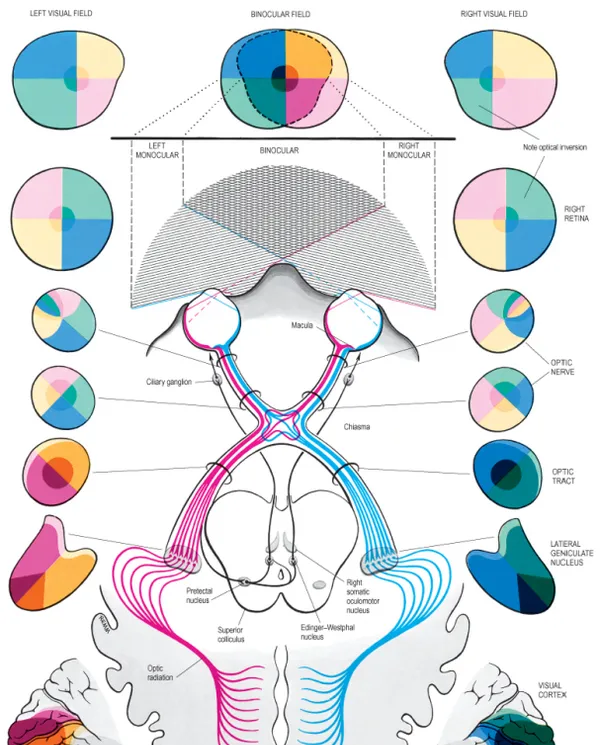
上圖是橫斷面的人腦視覺通路[1]示意圖。人眼的所有眼各有鼻側和顳側視野,在視網膜(Retina)感知光訊號之後,左右兩側的視神經將訊號向後傳遞,在視交叉(Optic Chiasma)將各自的鼻側和顳側訊號分流並繼續向後傳遞,分流之後左側神經只傳遞右側視覺訊號(左眼鼻側視野和右眼顳側視野),右側神經質傳遞左側視覺訊號(左眼顳側視野和右眼鼻側視野),分別透過兩側的外側膝狀體(LGN, Lateral Geniculate Nucleus),繼續傳遞到枕葉的初級視皮層(V1, Primary Visual Cortex)和更高級的視皮層。易於理解的,視網膜感知的物體,透過該通路,會以一定空間關系對映視皮層上[2], 該關系被稱為視網膜拓撲對映(Retinotopy)[3].
可以看到,人 腦的視覺訊號傳導通路,在進入大腦皮層之前是高度的結構化的、分工明確的。在進入初級視覺皮層之後,視覺訊號的處理會會表現出非常復雜的狀態 ,目前我們初級視覺皮層之後的視覺訊號處理的認識,並沒有徹底清楚,但是這並不妨害我們去認識人腦皮層對視覺訊號的認知模型,這便是下一部份的內容:視覺資訊處理的結構化特征。
1.2 視覺資訊處理的結構化特征
視覺訊號傳遞到初級視皮層V1之後,會繼續向更高級的視覺皮層傳遞,最早在根據獼猴(Macaque Monkey)視皮層的解剖結果,人們重建了其各個視皮層之間的分級結構[4]:
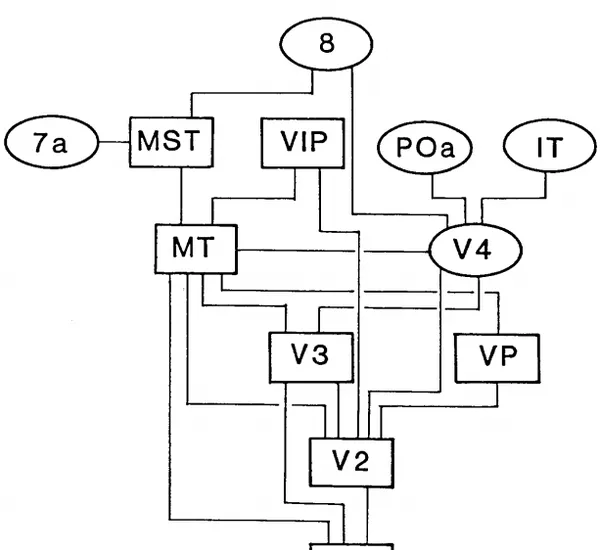
上圖中的縮寫表示的是人腦相關的皮層區域。以此為基礎,我們逐漸理解了人的視皮層分級結構:

從初級到更高級的視覺皮層,視覺資訊逐級傳遞。人腦理解的內容越來越復混成、抽象化,由「模式」變成具體的"物",再到物的特性和物與物之間的關系。
1.3 視覺認知的編碼特征
進入到初級視覺皮層的訊號是非常的繁雜和全面的,人腦並不需要所有的這些訊號,而是 只需要提取其中的有用訊號就可以了, 否則會給大腦的視覺理解帶來太大的壓力,基於1.2的結構化特征,人腦發展出了 視覺編碼壓縮方式 。
人腦會把透過視覺通路傳遞過來的訊號,跟自己的經驗比對,比照經驗中的常見影像模式,經過 比對之後,找到最為匹配的模式 (線、角、圓……;斑馬線,轉角……),並以該模式取代實際的事物。這就好比給視覺訊號做了一次編碼,以 人腦的內稟訊號取代實際訊號對視覺影像做了極大的壓縮 ,並為後續的快速處理做好了基礎。這種壓縮對應影片處理的幀內壓縮。
而對於運動影像,前後影像有很大的連續性,他們之間的相似度很高,可以 利用他們的相似性做進一步的壓縮 ,目前的影片處理演算法都利用了這一點,人腦自然也不會放過。這導致了視覺感知的視覺暫留現象。
總結來說,編碼的方式,主要有兩種:
以上都要求我們的 腦子快速的對影像進行理解,從而找到模式相似性和連續性特征 。
所以你感知的影像,並不是你真正眼睛看見的。
1.4 視覺和諧:一種可能的解釋
編碼的基礎在於對影像的理解,對於理解不了(無法快速找到基本模式)的事物影像,人腦就不能對其進行很好的壓縮,這會占用很大的精力。
先舉幾個例子
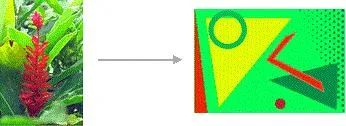
易於壓縮:



不易壓縮:


可以看出,易於壓縮的影像相比於不易壓縮的影像有以下幾個特點:
易於壓縮的影像,給人腦的資訊處理壓力低,不易壓縮的影像反之,會制造處理壓力,讓人有壓抑感和潛在的恐慌感。同時,你或許註意到,這些規律也可以套用到其他方面,比如:
視覺上的和諧,跟影像的視覺可壓縮性直接相關。
2 色彩的協調 (color harmony )
我們談論色彩的協調,實際上是在談論人的色覺的協調(color perception harmony)。基於上一部份的結論, 配色的協調對應的是較好的視覺可壓縮性 。那麽題中的問題就轉化為: 什麽樣的配色才具有較好的視覺可壓縮性 。我們從最基本的開始。
2.1 為什麽是RGB
我們都知道R(Red 紅色) G(Green 綠色) B(Blue 藍色)是三原色,他們不同成份的配合可以構成人眼所感知的任何顏色,那麽原因何在呢。
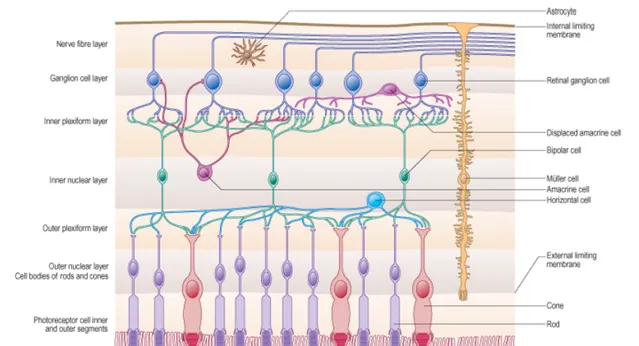
在人眼的視網膜上,主要有兩種感光細胞:視桿細胞(Rod)和視椎(Cone)細胞 [1]。視桿細胞有大概一億兩千萬個,在視網膜上廣泛分布,但是他們不能感知色彩,只感知光感,在光線不足的情況下,主要是視桿細胞作用,因此我們在晚上看不見色彩。視錐細胞則僅有7~8百萬個,主要分布在視網膜最為敏感的黃斑區(fovea)。並且對色彩敏感。按照對不同色彩(紅綠藍)的敏感性,可以進一步的劃分三種視錐細胞:短波(S)、中波(M)、長波(L)視錐細胞。
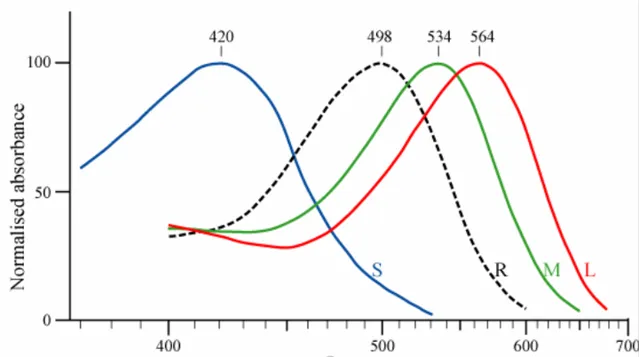
分別對藍色、綠色和紅色最為敏感。要註意各種視錐細胞也能感知其他色彩,只是對應的放電強度不如最為敏感的顏色。人能產生顏色的感覺,來自三種視錐細胞對不同波長光線的神經訊號差異, 人能感知的所有顏色可用這三組訊號的強弱表示 。也就是說 RGB色彩的混合,是人腦在視覺資訊處理的時候人工混合的。
三原色是人的色覺系統決定的,甚至不適用於其它動物(貓、狗等就有不同於人類的感光細胞)。 我們用三原色可以混合出其它顏色,是因為我們的視覺系統無法區分其他的顏色的神經訊號和三種神經訊號混合的訊號。 而物理上根本不存在顏色,不同顏色的光本質是不同波長的電磁波,顏色是視覺系統前進演化來的功能,以便於在大自然中辨識不同的事物。
2.2 色彩配合的可壓縮性
有了RGB基礎,我們可以定義在 視覺壓縮概念下的色彩相似性, 即相似顏色的組合,在讓視錐細胞產生的放電上模式上較為簡單, 盡量只有一種視錐細胞放電強,其他視錐細胞放電弱。 另一方面,我們還可以根據RGB的原理, 使主體顏色讓一種視錐細胞放電強,其他顏色讓其他視錐細胞放電強 ,這種情況下,能有效的突出主題,利於資訊壓縮。此外,還可以根據 模型匹配的原理,類似最為常見的自然的顏色模型,也是易於壓縮的。 以上三種情況分別對應著
- 相似色配色 (色彩接近)
- 互補色配色 (混合之後能產生白色)
- 擬自然配色 (粉色的花瓣配綠葉)
我們先根據RGB原理繪制一張色輪圖:
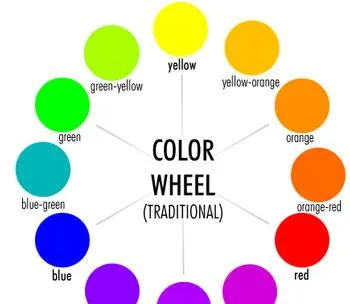
在色輪上,相似色位置鄰近:
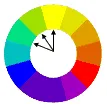
互補色則在對面:
擬自然配色則沒有在色輪上的規律:
有了以上的基礎,不僅能回答題主的問題,還能回答所有關於配色的問題。比如,對於題主問題的簡化回答是:
-
黑白灰的感知不需要色彩訊號的引入,視覺壓縮容易。
-
黑白灰配色是非常常見的自然界配色模式(比如在夜晚沒有顏色感知的時候),容易找到匹配的模式。
因此人腦在處理黑白灰配色上負擔小。再加上我們經常在莊重的場合看到黑白灰配色模式,我們在日常看到這種模式的時候,也會有嚴肅認真的感覺摻和進來(記憶因素的引入)。
以上
--------
[1] Standring, Susan. "Gray’s anatomy." The anatomical basis of clinical practice 39 (2008).
[2] Tootell R B H, Hadjikhani N K, Vanduffel W, et al. Functional analysis of primary visual cortex (V1) in humans[J]. Proceedings of the National Academy of Sciences, 1998, 95(3): 811-817.
[3] Engel S A, Glover G H, Wandell B A. Retinotopic organization in human visual cortex and the spatial precision of functional MRI[J]. Cerebral cortex, 1997, 7(2): 181-192.
[4] Van Essen, David C., and John HR Maunsell. "Hierarchical organization and functional streams in the visual cortex." Trends in neurosciences 6 (1983): 370-375.











