謝邀 @袁霖 ,回答這個問題之前,我們先來了解一下背景知識:
1. 華麗琴鳥( Menura novaehollandiae )
首先讓我們來認識一下華麗琴鳥,這個鳥屬於雀形目(Passeriformes)琴鳥科(Menuridae)。體長有80-100 cm,大概2斤重。身體為淺褐色,雄鳥擁有非常長的尾羽,有兩條金褐色的向內卷曲的外側尾羽,中間還有12枚微白色尾羽,極其漂亮。雌鳥顯得比較暗淡(BirdLife International, 2012)。

華麗琴鳥主要分布於澳洲東南部溫帶,熱帶雨林。主要食物為昆蟲和植物種子。華麗琴鳥被人所知還是其婉轉動聽的歌聲,而且非常善於模仿各種鳥類的鳴叫聲,還能學人間的各種聲音。如汽車喇叭聲等,也是澳洲鳥類中最受人喜愛的珍禽之一(Christidis and Boles, 2008)。
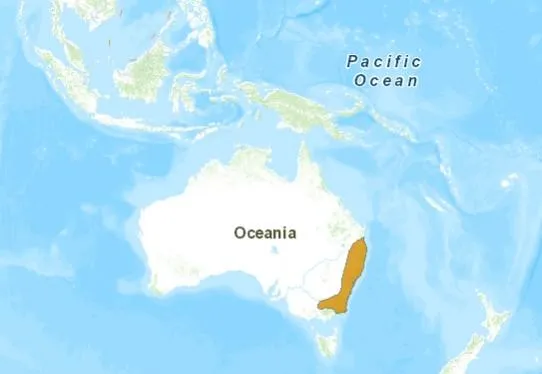
華麗琴鳥的分布(圖:IUCN)
了解了華麗琴鳥之後,再給普及一下鳥類的鳴聲的背景知識。
2 鳥類的鳴聲
2.1 鳥類的發聲器官
2.1.1 鳴管
鳴管是鳥類特有的發聲器官,位氣管下端與支氣管交界處。但是也有少數鳥類沒有鳴管(鴕鳥,鸛,這些鳥會透過其他方式發聲,比如鸛用其上下嘴急速啪打,發出「嗒嗒嗒」的嘴響聲)。
有些鳥透過鳴管內外兩側的鳴膜,隨著氣流的震動發出聲音。很多善於鳴叫的鳥類,除了鳴膜,還有半月膜(沿著鳴骨有一正中的薄膜伸出),透過氣流進出半月膜,振動發出聲音。鳴骨下方有氣室,可以調節氣流和壓力,調節不同的聲音。
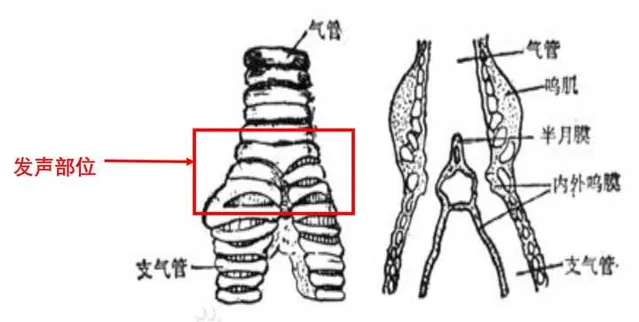
圖片來自百度百科
2.1.2 鳴肌
鳴肌是控制鳴管管徑、曲率以及鳴膜張力的特殊肌群。鳴肌受到神經的支配,可以發出不同頻率,婉轉多變的聲音,鳴肌的復雜程度和鳥類的鳴叫密切相關。比如低等的非雀形目鳥類大多只有簡單的鳴肌,甚至沒有鳴肌(鸛,信天翁)。鴨類,鴿類以及多數雞形目只有一對鳴肌,蜂鳥和低等雀形目鳥類有2對鳴肌,鸚鵡類有3對鳴肌,有些小型鳴禽有5-9對(通常6對)(Gill, 2007)。
2.2 鳥類的發聲機理
鳥類的發聲涉及聲音的產生和共鳴兩個方面:氣流進入鳥類氣囊與肺部,流經鳴管時,使得鳴膜受到振動而發出聲音( 聲音的產生 ),鳴膜在鳴肌的調節下,緊繃程度有所不同,從而產生不同的諧波( 共鳴 ),構成多變的聲調(Gill, 2007)。
2.3 鳥類鳴聲的分類
鳥類的鳴聲可以分成兩類,一類是鳴叫,指的是短促,簡單的鳴聲;另外一類是鳴唱,指的是雄鳥在繁殖期內發出的較長的,相對復雜的鳴聲。
2.4 鳥類鳴聲的功能和作用
2.4.1 鳴叫: 鳥類的鳴叫功能比較多,透過鳴叫可以進行胚胎之間的交流,親子之間的辨識,集群之間的通訊,雛鳥乞求食物,天敵的警戒等(鄭光美, 2012)。

加拿大雁( Branta canadensis )透過鳴叫可以和自己的孩子交流(圖:小葉叔叔)
2.4.2 鳴唱: 雄鳥繁殖期間的鳴唱主要有兩個功能,一個是保衛領域,另一個是建立並維持配偶關系(找物件)(鄭光美, 2012)。

黃鸝( Oriolus chinensis )正在鳴唱,宣布自己領域或者尋找物件,圖片來源鳥網
華麗琴鳥為什麽能模仿幾乎一切它聽到的聲音呢?
鳥類的效鳴
鳥類模仿其他種類鳴聲的現象呢,我們專業術語叫 效鳴(mimicry) 。例如我們熟悉的八哥,百靈鳥都能模仿多種動物的叫聲,已發現15-20%的鳴禽具有效鳴的行為。有些鳥兒除了可以模仿其他鳥類的叫聲之外,還可以模仿青蛙,昆蟲,貓等動物的叫聲,甚至模仿汽笛聲,人說話的聲音(Zann and Dunstan, 2008)。
關於鳥類效鳴的生物學意義,目前尚存爭議,主要有以下三個假說:
錯誤學習假說( learning mistakes hypothesis ): Hindmarsh(1986)透過紫翅椋鳥( Sturnus vulgaris )的研究發現,雄鳥的效鳴並沒有對性選擇有幫助,所以作者認為效鳴是鳥類鳴聲學習錯誤導致的,其生存環境中其他種類的叫聲影響到模仿者的學習行為,無意中學習到,效鳴現象是鳥類聲音學習過程中沒有任何意義的產物。Garamszegi et al. ,(2007)透過歐洲241種雀形目鳥類的效鳴研究也證實了錯誤學習假說;
資源競爭假說( resource competition hypothesis ): Krebs (1977)和 Rechten(1978)認為認為鳥類可以透過效鳴模仿周圍其他同種或者不同種鳥類的鳴聲,讓其他鳥類誤以為這個區域存在激烈的種內競爭,讓使對手放棄進入這個區域,也就是資源競爭假說(Baptista and Catchpole, 1989)。這個假說後來也在大山雀( Parus major )(Gorissen et al., 2006),黃冠鸚哥( Amazona ochrocephala )(Wright et al., 2005)等鳥類中得到證實;
性選擇假說( sex selection hypothesis ): 目前更多的學者認同Hartshorne(1956)提出性選擇假說,這個假說認為效鳴是雄鳥在性選擇過程中擴大曲目訊號的一種方式,用來吸引更多的雌鳥,復雜的音節也對雌鳥更有吸重力(Ferguson et al. , 2002),提高吸引雌性成功率(Colemanet et al., 2007),進而提高適合度。
根據學者研究發現 (Zann and Dunstan, 2008),華麗琴鳥可以模仿30多種其他鳥類的鳴聲。這裏我總結 華麗琴鳥能模仿這些聲音的幾個原因:
1. 特殊的生理結構
從背景知識我們了解到,鳥兒的歌唱能力主要取決於發達的鳴肌,華麗琴鳥擁有3對鳴肌,使其擁有較強的聲音模仿能力(Kaplan,1999)。
2. 性選擇壓力
研究發現,雄性的華麗琴鳥效鳴能力極其強,雖然雌鳥也會效鳴,但是鳴聲較少,其歌聲和效鳴多用於保衛雛鳥,防禦天敵和種內競爭。雌鳥自己唱歌少,但是卻偏愛歌唱家,喜歡選擇那些曲目種類多而且音樂轉換多的雄性個體。那些唱得少,且不好聽的雄性個體往往找不到物件,不能繁殖後代,久而久之剩下的個個都是歌唱家(Dalziell and Welbergen, 2016)。
3. 後天 勤於學習
研究還發現,不同年齡段的華麗琴鳥的歌唱曲目是不同的,年幼的個體明顯曲目唱得少,這些效鳴多樣性並不是天生的,都是後天學習的(Zann and Dunstan, 2008)。
實際上,大部份鳥類模仿其他鳥類的鳴聲只是占了它自己所有鳴聲中的一小部份,而且它只能模仿其他種類鳴聲的一些片段。由於遺傳和生理的差異,效鳴的聲音還達不到原來個體鳴唱的音色。但是最近透過研究發現華麗琴鳥的模仿能力極其強,模仿的灰鵙鶇( Colluricincla harmonica )聲音可以讓灰鵙鶇有所察覺,作出靠近的反應,可見 華麗琴鳥技藝高超 (Dalziell and Magrath, 2012)。

灰鵙鶇(圖:
http://www. indonesiatravelingguide.com /indonesia-bird/colluricincla-harmonica/)
參考文獻:
鄭光美. 鳥類學(第二版). 北京師範大學出版社. 2012
BirdLife International. Menura novaehollandiae. The IUCN Red List of Threatened Species 2012: e.T22703605A39248049.2012.
Coleman S W, Patricelli G L, Coyle B, et al. Female preferences drive the evolution of mimetic accuracy in male sexual displays. Biology Letters, 2007, 3(5): 463-466.
Christidis, L. and Boles, W.E. Systematics and taxonomy of Australian birds. CSIRO Publishing, Collingwood, Australia. 2008.
Dalziell A H, Magrath R D. Fooling the experts: accurate vocal mimicry in the song of the superb lyrebird, Menura novaehollandiae . Animal Behaviour, 2012, 83(6): 1401-1410.
Dalziell A H, Welbergen J A. Elaborate Mimetic Vocal Displays by Female Superb Lyrebirds[J]. Frontiers in Ecology and Evolution, 2016, 4: 34.
Garamszegi L Z, Eens M, Pavlova D Z, et al. A comparative study of the function of heterospecific vocal mimicry in European passerines. Behavioral Ecology, 2007, 18(6): 1001-1009.
Gill F B. Ornithology(Third edition). W.H.Freeman and Company, Nwe York, 2007.
Gorissen L, Gorissen M, Eens M. Heterospecific song matching in two closely related songbirds ( Parus major and P. caeruleus ): great tits match blue tits but not vice versa. Behavioral Ecology and Sociobiology, 2006, 60(2): 260-269.
Ferguson J W H, Van Zyl A, Delport K. Vocal
mimicry in African Cossypha robin chats. Journal für Ornithologie, 2002,
143(3): 319-330.
Hartshorne C. The monotony-threshold in singing birds. The Auk, 1956: 176-192.
Hindmarsh A M. The functional significance of vocal mimicry in song. Behaviour, 1986, 99(1): 87-100.
Kaplan G. Song Structure and Function of Mimicry in the Australian Magpie (
Gymnorhina tibicen
): Compared to Lyrebird (
Menura ssp
.). International Journal of Comparative Psychology, 1999, 12(4).
Krebs J R. The significance of song repertoires: the Beau Geste hypothesis. Animal Behaviour, 1977, 25: 475-478.
Rechten C. Interspecific mimicry in birdsong: does the Beau Geste hypothesis apply?. Animal Behaviour, 1978, 26: 305-306.
Wright T F, Rodriguez A M, Fleischer R C. Vocal dialects, sex-biased dispersal, and microsatellite population structure in the parrot Amazona auropalliata . Molecular Ecology, 2005, 14(4): 1197-1205.
Zann R, Dunstan E. Mimetic song in superb lyrebirds: species mimicked and mimetic accuracy in different populations and age classes. Animal Behaviour, 2008, 76(3): 1043-1054.











