上一次给大家介绍了表观遗传修饰中的DNA甲基化修饰,今天给大家介绍表观遗传的另一大修饰组蛋白甲基化。
核小体是由核心组蛋白(Histone, H)八聚体(pA × 2、pB × 2、p ×2、p × 2)与缠绕其外周长度为146bpDNA组成的核心颗粒及颗粒之间50bpDNA和一个p构成。
组蛋白能维持DNA结构、保护遗传信息和调控基因表达。组蛋白氨基末端(N端)结构域伸出核小体,可同其他调节蛋白和DNA发生相互作用。组蛋白修饰有甲基化、磷酸化、乙酰化、巴豆酰化、泛素化、糖基化、ADP核糖基化等。组蛋白修饰失衡可导致肿瘤发生发展,且组蛋白p和p残基甲基化和乙酰化的丧失已被证实是肿瘤细胞的标志物。
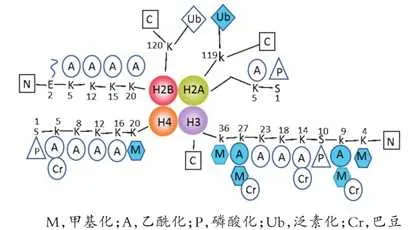
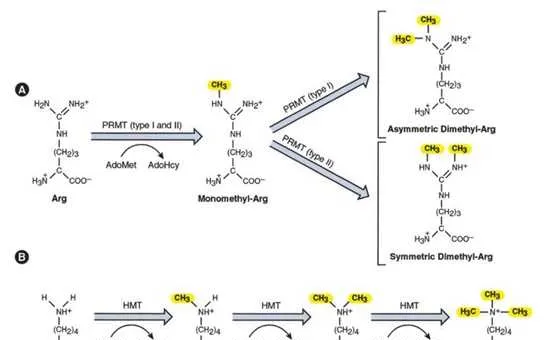
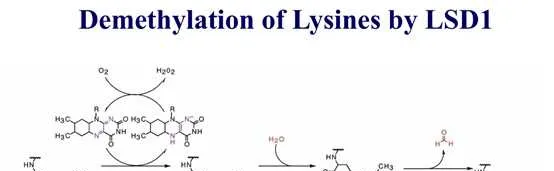
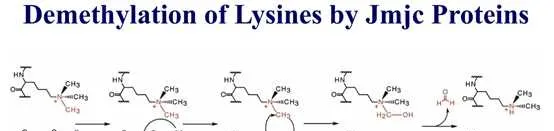
组蛋白甲基化(histone methylation)是发生在p和p组蛋白N端赖氨酸(K)或精氨酸(S)残基上的甲基化。其功能旨在形成和维持异染色质的结构、基因组印迹、DNA修复、X染色质的失活和转录等调控方面。组蛋白甲基化过程主要由组蛋白甲基转移酶(histone methyltransferase,HMT)催化,而HMT又可分为组蛋白赖氨酸甲基转移酶(histone lysine methyltransferase,HKMT)和组蛋白精氨酸甲基转移酶(protein arginine methyltransferase,PRMT)。而组蛋白去甲基化酶,大致分为LSD(Lysine-specific demethylase)和JMJD(JmjC domain-containing family)两个家族。LSD1能特异性地去除组蛋白pK4和pK9的单双甲基化修饰,而Jmic 家族能去除赖氨酸三甲基化的修饰。p赖氨酸(pK)4、9、27、36、79和p赖氨酸(pK)的20位点可被甲基化,其中组蛋白pK4和pK9是常见的两个修饰位点。赖氨酸残基可以发生单、双或三甲基化修饰,而精氨酸残基则只发生单和双甲基化修饰(对称或不对称)。
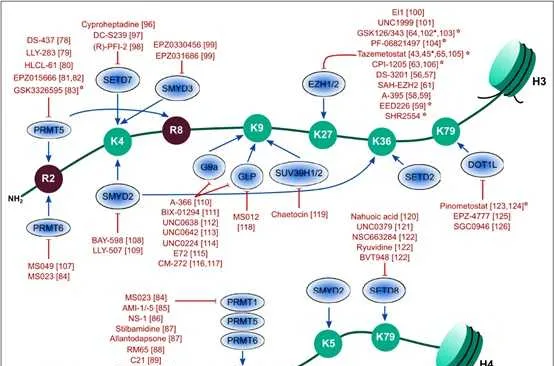
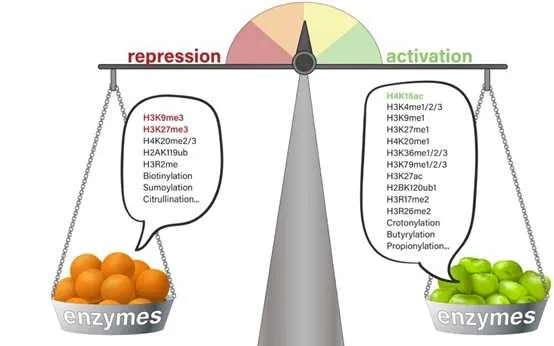
一般来说,组蛋白p和p不同位点的甲基化以及甲基化数量对基因的转录调控具有很大的意义。其中pK9me3,pK27me3以及pK20me2/3介导的是转录抑制,而pK4me1/2/3、pK9me1、pK27me1、pK36me1/2/3、pk79me1/2/3介导的则是转录激活。更多的修饰位点所介导的转录活性如下图所示。
1 组蛋白甲基化修饰酶
赖氨酸可以被六种主要的组蛋白赖氨酸甲基转移酶复合物(KMT1-6)单、二或三甲基化。KMT1家族在哺乳动物中至少包含4个成员,包括SUV39p/2、G9a、GLP和SETDB1,并以pK9为甲基化底物。KMT2家族酶存在于大分子复合物中,称为与Set1相关的蛋白质复合物(COMPASS),并在pK4上进行单甲基、二甲基或三甲基标记。KMT3家族包括NSD1、NSD2 (WHSC1)和NSD3 (WHSC1L1),并主要甲基化pK36。KMT4家族的唯一成员是DOT1L,它实现了pK79的甲基化。KMT5家族包括PR- Set7和SUV4-20p/2,它们分别实现pK20单甲基化和二/三甲基化。KMT6家族包括用于pK27单甲基化、二甲基化和三甲基化的功能冗余酶EZp和EZp。
自赖氨酸去甲基化酶LSD1被发现以来,赖氨酸甲基化一直是一个可逆过程。至少有6个家族的组蛋白赖氨酸去甲基化酶具有独特的和重叠的功能。KDM1家族包括LSD1 (KDM1A)和LSD2 (KDM1B),两者都能使pK4me2/me1脱甲基,但不能使pK4me3脱甲基。此外,LSD1还可以通过将其与CoREST相互作用的抑制复合物转换为与雄激素受体(AR)相互作用的激活复合物来参与pK9的去甲基化。与LSD家族不同,赖氨酸去甲基酶的所有其他家族成员都含有Jumonji (JmjC)结构域,由于涉及的化学成分不同,该结构域有潜力去除三甲基标记。JHDM1A (KDM2A)和JHDM1B (KDM2B)属于KDM2家族,对pK36me2/me1和pK4me3活性较高 。JHDM1A是第一个鉴定出的含有JmjC结构域的去甲基酶。KDM3家族包括KDM3A、KDM3B和JMJD1C,对pK9me2/me1具有去甲基化酶活性。KDM4家族包括KDM4A、KDM4B、KDM4C和KDM4D,对pK9me3/me2和pK36me3/me2具有不同的去甲基化酶活性。KDM5家族包括KDM5A、KDM5B、KDM5C和KDM5D,它们都能对pK4me3/me2进行脱甲基。KDM6家族包括UTX (KDM6A)、JMJD3 (KDM6B)和UTY。UTX和JMJD3对pK27me3/me2有特异性,而Y-连接的类似物UTY催化活性较低。一些KDM被认为是多种癌症发展的促进因素,因此被认为是潜在的药物靶点。
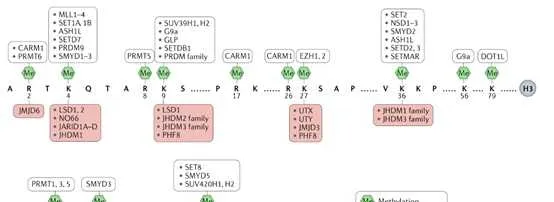
2 组蛋白赖氨酸不同位点的甲基化修饰
组蛋白pK4的甲基化是由COMPASS家族中的SET1A、SET1B和MLL1-4在内的增强子和启动子实现的。COMPASS的不同亚基也被证明可以调控pK4的二甲基和/或三甲基化,包括WDR5、AspL、RbBP5和Dyp30,这些亚基是COMPASS家族所有成员共有的。有趣的是,SET1A的致癌功能已经通过组蛋白和非组蛋白底物YAP的甲基化分别与乳腺癌转移、肺癌和结直肠癌的肿瘤发生有关。MLL1和MLL2在启动子和/或Polycomb响应元件(PRE)上实现二甲基化和三甲基化,MLL2也可以在二价基因的启动子和增强子上实现pK4的甲基化。MLL3和MLL4能够在增强子处使pK4单甲基化。在急性髓系和淋巴系白血病(AML和ALL)中,MLL1经常通过与其他致癌基因易位而发生突变,在儿童白血病中约占80%,在成人白血病中占5% 10%。嵌合蛋白缺乏MLL1的催化SET结构域,并驱动白血病发生。最近,我们确定了治疗MLL重排白血病的策略,通过稳定MLL的野生型拷贝来减弱由MLL融合蛋白及其致癌辅助因子超延伸复合物(SEC)介导的异常转录。MLL3和MLL4也在癌症中均被发现高度突变。组蛋白pK4me3标记可以帮助招募染色质重塑因子CHD1和BPTF,这两种染色质重塑因子可以帮助染色质打开。
由于SET2酶与RNA Pol II的CTD磷酸化形式相关联,在积极转录的基因体内检测到组蛋白pK36me3。由ASpL和NSD1-3家族实现的pK36me2的功能不太为人所知。最近,一个潜在的pK4me3和pK36me2之间的串扰在LEDGF的中心发生。LEDGF通过其整合酶结合域直接与Menin和MLL1相互作用,是MLL1依赖的转录和白血病转化所必需的。同时,LEDGF通过其PWWP结构域与二甲基化的pK36结合。LEDGF引起了越来越多的关注,因为研究表明LEDGF在MLL重排白血病中至关重要,而不是在造血系统中,这提高了有效靶向LEDGF而不产生一般副作用的治疗潜力。通过使用CP65(一种用于抑制HIV病毒复制的环状肽)靶向LEDGF,治疗mll重组白血病取得了有限的成功,因为LEDGF上的相同结构域结合了HIV整合酶和MLL1。利用蛋白裂解靶向嵌合(PROTAC)技术降解LEDGF可能是一个新的方向。pK36 me3也可以阻止同一组蛋白尾部PRC2介导的pK27残基的甲基化。
由DOT1L实现的组蛋白pK79甲基化标记,位于与活性基因表达相关的组蛋白的球状结构域。DOT1L也是唯一一种催化赖氨酸甲基化的酶,它与具有SET结构域甲基转移酶不同,而pK79的去甲基化酶至今尚未被鉴定出来。DOT1L是在一个名为DotCom的复杂结构中发现的,它与MLL转移伙伴AF9或其同源物ENL和AF10一起。DOT1L活性还可促进乳腺癌细胞增殖和转移。白血病中pK79甲基化异常上调导致了DOT1L抑制剂EPZ- 5676的开发和使用,用于治疗MLL重排白血病,目前正在临床研究中。
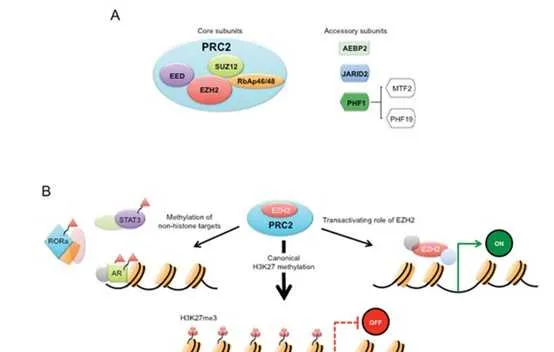
组蛋白pK9和pK27甲基化是形成不同形式的异染色质所必需的。组蛋白pK9me3和pK27me3被认为是唯一真正的表观遗传标记,因为它们已经定义了在DNA复制后可遗传的机制。HP1蛋白HP1α (CBX5)、HP1β (CBX1)和HP1γ (CBX3)含有甲基赖氨酸结合的染色体结构域,在异染色质形成中发挥重要作用。SUV39p催化的组蛋白pK9的甲基化为HP1蛋白创造了一个结合位点,而HP1蛋白又会吸收更多的SUV39p,这一机制有助于异染色质形成的增殖。在pK27me3的形成中,EZp在PRC2复合物中实现pK27甲基化,而EED亚基识别这种甲基化,并变构性地进一步激活EZp的SET结构域。与COMPASS家族催化的pK4me1/2/3的独特分布类似,pK27me1/2/3的分布在整个基因组中是相互排斥的,pK27me3主要位于启动子(特别是二价基因),pK27me2主要位于基因间区域,pK27me1主要位于转录活性基因的基因体中。由于PRC2的EZp和SUZ12亚基对于HP1α的稳定是必需的,因此异染色质标记pK27me3和pK9甲基化可能协同维持染色质上的HP1α,突出pK9me2/3和pK27me3基因沉默途径之间的关键交叉。当EZp在癌细胞中异常表达或突变时,EZp抑制剂常用于防止肿瘤抑制基因的组蛋白甲基化。
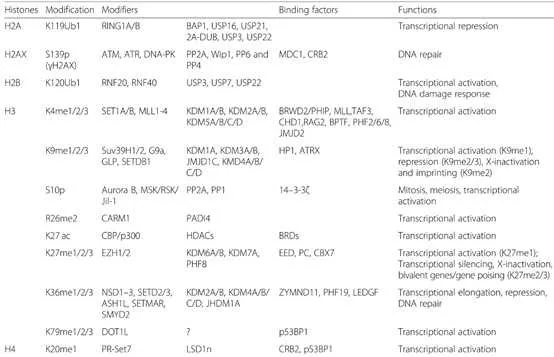
3 组蛋白甲基化与癌症
一些证据表明异常组蛋白甲基化可能在癌症中发挥作用。最初的研究表明,某些组蛋白甲基化水平的变化与癌症复发增加和低生存率相关。尽管这些变化是否具有因果关系尚有待确定,然而,它们可能发展成为潜在的生物标记物,用于药物发现和诊断或预后。最近的研究提供了越来越多的遗传学证据,表明组蛋白甲基化事件在肿瘤发生中起着因果作用。组蛋白甲基修饰和甲基结合蛋白的突变或表达改变与各种不同癌症的发病率增加相关。例如,pK27me3甲基转移酶在一些癌症中上调,包括前列腺癌、乳腺癌和淋巴瘤。重要的是,最近发现EZp的激活点突变与B细胞淋巴瘤相关,这与EZp致癌的观点一致。与此一致的是,在多种人类癌症中发现了pK27去甲基化酶UTX的体细胞失活点突变。然而,EZp并不总是作为致癌基因;在骨髓增生综合征中发现了导致EZp甲基转移酶活性丧失的突变,这表明EZp在该癌症类型中起着抑癌作用。EZp作为癌基因或抑癌基因的双重作用突出了癌基因和抑癌基因的背景依赖性,并提出了pK27me3在不同细胞类型中可能具有不同功能的可能性。
D-2-羟基戊二酸酯(D2HG)是一种肿瘤代谢物,可抑制许多去甲基化酶,导致基因组和转录组甲基化谱的改变,以及基因表达和基因组拓扑结构的改变。肿瘤发生与编码异柠檬酸脱氢酶1 (IDp)和IDp的基因的特定突变有关,这些突变阻止异柠檬酸转化为α-酮戊二酸,并促进α-酮戊二酸还原为其结构类似物D2HG。参与组蛋白去甲基化酶的Jumonji C结构域家族都是铁依赖双加氧酶,它们在染色质上的催化活性被高水平的D2HG竞争性地抑制。

4 组蛋白甲基化靶向治疗——组蛋白甲基化调节剂
在过去的几年里,已经开发了大量不同组蛋白甲基转移酶的特异性抑制剂。EZp、DOT1L、PRMT1、PRMT5抑制剂目前已进入临床试验阶段,其中EZp抑制剂的临床开发最为先进。并且在2020年1月23日,FDA宣布全球首个EZp抑制剂Tazemetostat重磅上市,批准的适应症为不适用于手术切除的转移性或晚期的上皮样肉瘤。
EZpi: EZp是PRC2的催化亚基;其他PRC2亚基包括zeste 12抑制子(SUZ12)、胚胎外胚层发育蛋白(EED)和视网膜母细胞瘤结合蛋白p46(也称为RBBP7)。EZp通过催化pK27的单甲基化、二甲基化和三甲基化来促进基因沉默。EZp同源物EZp具有较弱的组蛋白甲基转移酶活性,但在特定情况下,特别是当EZp水平较低时,仍可能参与pK27甲基化。EZp对B细胞成熟至关重要,是多发性骨髓瘤、滤泡淋巴瘤和弥漫大B细胞淋巴瘤(DLBCL)有前景的治疗靶点。约20% - 30%的滤泡性淋巴瘤和生发中心DLBCLs在EZp SET域的特定残基中存在杂合点突变(Y641, A677, A687),它增强了二甲基化和单甲基化的靶向性,从而导致pK27me3水平的大幅提高。在BRCA1相关蛋白1 (BAP1)突变的恶性间皮瘤和染色质重塑复合物SWI/SNF缺陷的肿瘤中也可以看到对EZp活性的获得性依赖,例如,在SWI/SNF亚基SMARCB1缺失的恶性横纹肌肿瘤中。
SWI/SNF核小体重塑复合物可拮抗PRC2介导的基因沉默,并已被证明可从染色质中驱逐Polycomb因子。SWI/SNF失活与EZp突变在一系列癌症中合成致死。虽然SWI/SNF基因突变被认为通过获得PRC2功能和沉默肿瘤抑制基因来驱动转化,但EZp的非催化活性也被认为与肿瘤发生有关。有研究表明,SWI/SNF和PRC2功能的联合缺失可能导致细胞死亡,这是由于整体转录放松,而不是特定PRC2靶点的去抑制。BAP1是一种去泛素化酶,靶向泛素化的pAK119,从而反对Polycomb介导的基因沉默;然而,BAP1丢失后对EZp抑制剂的敏感性似乎是细胞类型特异性的,在间皮瘤中观察到,但在葡萄膜黑素瘤中未观察到。PRC2还可以作为肿瘤抑制因子,在骨髓增生异常综合征和慢性骨髓增生性肿瘤中有报道称EZp反复失活突变。编码其他核心PRC2成分的基因EED和SUZ12的功能缺失突变也见于T细胞急性淋巴细胞白血病和恶性周围神经鞘肿瘤。PRC2在不同肿瘤环境中的功能不同被认为反映了特定的细胞转录程序和染色质环境在确定PRC2靶向基因方面的关键作用,并强调了仔细监测EZp抑制剂治疗的患者的必要性。
EZp异常激活是多种癌症的一个特征,包括乳腺癌、去雄前列腺癌、小细胞肺癌(SCLC)和神经母细胞瘤,并已被证实与肿瘤发生和获得干细胞样转录程序有关。尽管在某些癌症中,EZp的上调可能是恶性过程的结果而不是驱动因素139,但临床前研究表明,在许多这些肿瘤类型中,EZp的缺失或抑制会损害增殖和肿瘤生长。多种EZp抑制剂目前正在一系列癌症的I/II期临床试验中进行评估。令人鼓舞的是,在滤泡性淋巴瘤中,71%的EZp SET域突变激活的患者对EPZ-6438(他泽美司他)有应答,11%的患者达到完全应答。相比之下,没有EZp突变的患者只有33%的应答,31%的患者病情进展。
其他抑制剂:其他小分子KMT抑制剂已经被开发为潜在的抗癌治疗药物,包括靶向pK9 KMT的化合物。肺癌细胞系中pK9特异性KMT G9a的表达增加,使用抑制剂BIX-01294可以减少pK9的甲基化。类似地,SETDB1在黑色素瘤和肺癌中经常被放大,mithramycin治疗会下调SETDB1以抑制增殖。此外,天然产物毛霉素被鉴定为SUV39H的抑制剂,SUV39H是一种KMT,调节红细胞和B细胞分化。
PRMTs在癌症中经常过度表达,因此也成为了有吸引力的抗癌策略靶点。PRMT家族有9个成员(PRMT1-9)将精氨酸甲基化形成甲基精氨酸。I型PRMTs (PRMT1-6和8)催化不对称二甲基精氨酸的形成,而2型PRMTs (PRMT5和9)催化对称二甲基精氨酸的形成。组蛋白精氨酸甲基化标记可以激活(pR3me2a, pR2me2s, pR17me2a, pR26me2a)或抑制(pR2me2a, pR8me2a, pR8me2s, pR3me2s)基因转录,并且在多种癌症中已经观察到PRMT功能障碍。在NSCLC中有过表达PRMT1和PRMT4 (CARM1)的报道,在乳腺癌中PRMT4可以通过解除SWI/SNF的调控来驱动c-Myc通路的表达。在淋巴瘤、白血病、胶质母细胞瘤和前列腺癌中观察到PRMT5表达增加,据报道这些肿瘤激活了c-Myc和其他致癌转录因子。此外,有报道称PRMT7可抑制E-cadherin的表达,促进EMT在乳腺癌中的表达。临床试验中的PRMT抑制剂是GSK3326595(原名EPZ015938)和JNJ-64619178,目前正在对实体瘤和非霍奇金淋巴瘤患者进行评估。一些抑制剂已在临床前研究中显示出前景,如抑制AML模型中MLL-GAS7或MOZ-TIF2融合转化功能的PRMT1抑制剂AMI-408,以及在套细胞淋巴瘤异种移植物中显示抗肿瘤活性的PRMT5抑制剂EPZ015666。
组蛋白去甲基化酶LSD1 (KDM1A)在几种癌症中高表达,并且是造血细胞最终分化所必需的。LSD1通常使pK4me1/2去甲基化,从而抑制转录,但当LSD1与雄激素受体相互作用时,其酶活性切换到pK9me1/2,从而刺激转录(98)。LSD1也是G9a KMT的底物,在雄激素依赖的基因表达过程中,LSD1的甲基化会刺激CDH的募集。LSD1有一个C端类胺氧化酶结构域,在结构上与单氨氧化酶(MAO)相关。因此,MAO抑制剂tranylcypromine (TCP)可抑制LSD1,但其使用受到任意抗MAO活性的限制。一些更有选择性的TCP衍生物已经开发出来,其中一些已经进入了临床试验。在异种移植中,ORY-1001已被证明可以减少pK4me2和LSD1靶基因的表达,并减少肿瘤的生长。GSK2879552促进AML细胞分化并抑制其增殖。此外,最近有报道称LSD1抑制剂GSK354和GSK690在体外抑制细胞生长。有趣的是,小分子HDAC抑制剂4SC-202具有抑制LSD1的双重功能,具有相似的效力,目前正处于晚期血液系统恶性肿瘤患者的临床试验中。
虽然有四种靶向LSD1的化合物正在进行临床试验,但含有去甲基酶活性的JmjC结构域家族蛋白抑制剂的开发一直比较困难。其中两种化合物是GSK-J1及其前体药物GSK-J4,它们能抑制KDM6A (UTX)和KDM6B (JMJD3),但对KDM5A和5B的活性较低。这些蛋白质都与癌症有关。例如,KDM6A失活突变已在AML、多发性骨髓瘤和膀胱癌中报道,而KDM6B在TALL和转移性前列腺癌中高表达。此外,在10%的儿童急性巨核细胞白血病中发现KDM5A (JARID1) Nup98融合蛋白。最近,一种KDM5B的强效选择性抑制剂EPT1013182已被报道在细胞系中具有抗增殖作用,并在异种多发性骨髓瘤模型中抑制生长。